
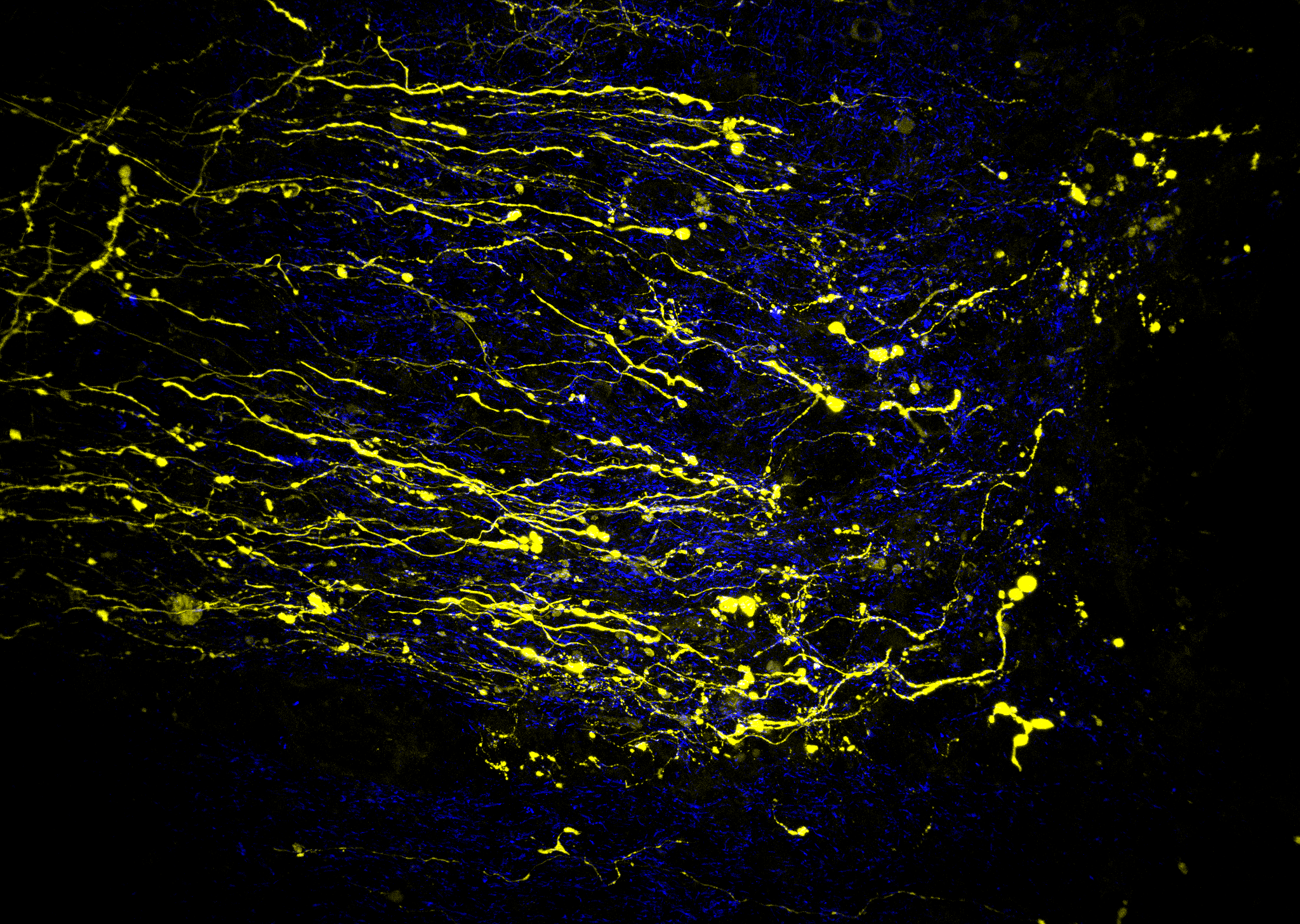

Molecular and Cellular Physiology of Synapse Formation
Research Focus
We’re interested in elucidating the molecular recognition events between pre- and postsynaptic nerve cells during synapse formation. These events are critical to nervous system function allowing the system to wire itself correctly during development. We wish to know how the presynaptic axon identifies the correct target cell, then how its neurosecretory apparatus becomes correctly aligned with the appropriate neurotransmitter receptors on the postsynaptic cell. To understand these events we are first identifing the proteins that mediate them, the receptors of the presynaptic plasma membrane and their postsynaptic ligands. Our particular focus recently has been on the neuromuscular synapse and the closely-related synapse from the electric organ of the marine electric ray. Using biochemical and molecular biological methods, we have found several potential receptors and ligands.
Research Details
The presynaptic calcium channel acts as a receptor for a synaptic laminin. That is, on the nerve terminal surface we have found the alpha 1 subunit of the calcium channel bound to a synapse-specific extracellular matrix protein, α4β2γ1 laminin, in the synaptic cleft. The c-terminal domain of the β2 laminin chain binds the 11th extracellular loop of the calcium channel. In addition, this calcium channel is also linked to the cytoskeletal protein spectrin in the nerve terminal cytosol. Spectrin is thought to be involved in attaching synaptic vesicle clusters to sites of neurotransmitter release. Thus, we hypothesize that the calcium channel may be a central presynaptic membrane component linking extracellular α4β2γ1 laminin with the neurosecretory apparatus. By secreting this laminin into the synaptic cleft, the postsynaptic cell could position the neurosecretory apparatus in the directly adjacent nerve terminal membrane.
To test our hypothesis, we have disrupted in vivo the interaction between the β2 chain of laminin and the nerve terminal calcium channel at the neuromuscular synapse. We exposed postnatal mouse muscle to a soluble fusion protein containing the 11th extracallular of the nerve terminal calcium channel. Our expectation was that this loop would occupy the calcium channel binding site on the β2 laminin chain, preventing the laminin from attaching to the calcium channel, and disassembling neurotransmitter release sites. This is exactly what occurs.
We have also found on the nerve terminal surface a second laminin composed of an α5, a β1 and a novel γ chain. This laminin is complexed with a transmembrane synaptic vesicle proteoglycan, SV2. The SV2/laminin complex is exclusive to the presynaptic plasma membrane and is not found with SV2 in the synaptic vesicle. Our data suggest that the SV2/laminin complex functions as an anchor for endocytosis zones.
Conclusion
Thus, our data supports an outside-in model for the differentiation of the neuronal plasma membrane. The extracellular matrix secreted by one cell determines the arrangement of the cellular machinery of its neighbor. At the presynaptic membrane the topological arrangement of both exocytotic and endocytic zones may be determined by the synapse specific laminins in the basal lamina. These laminins are deposited in the synaptic cleft by the muscle cell, presumably in response to basal lamina proteins like agrin or aria secreted by the nerve terminal. The extracellular matrix may be the medium through which neighboring cells establish long-term communication.